Воспользуйся поиском! Ты найдешь ответы на 99% своих вопросов.
Навигация
Сейчас на сайте
Просто реклама
Обособленность колонн
Чтобы сформировались колонны, необходимо несколько определенных условий.
Во-первых, должна быть определенная численность колонны. Например, семья рыжих
лесных муравьев не может быть слишком малой численности, так как в этом случае
она оказывается нежизнеспособной. Минимум численности жизнеспособной семьи
где-то около 100 тыс. Гнездо семьи из 100 тыс. муравьев имеет диаметр около 60
см и высоту 40-45 см. Такой муравейник еще слишком мал, чтобы иметь колонны (они
появляются позже при численности семей 180-200 тыс. особей). Число колонн в
муравейнике невелико: две, реже три (60-100 тыс. муравьев в одной колонне).
Второе необходимое условие - достаточно выраженная разобщенность колонн,
прежде всего пространственная. Без этого колонны существовать не могут.
Разобщение необходимо и в гнезде, и на кормовом участке. Это установлено и
проверено на разных видах муравьев. В основе обособленности колонн лежит
характерный для муравьев принцип вторичного деления территории*, выявленный в
1956 г. польским мирмекологом Я.Добжанской. Каждый охотник действует на
элементарном поисковом участке, с которым он связан длительное время. Если он и
меняет участок, то это происходит только в пределах одного из секторов
охраняемой территории семьи, точнее - в пределах одной дороги. Этим обусловлено
постоянство сочетаний рабочих муравьев на кормовых дорогах, по которым муравьи
попадают к своим поисковым участкам и колониям тлей. Сами дороги также
отличаются большим постоянством, оставаясь практически неизменными 10-13 лет.
Эти цифры означают лишь максимальные сроки регулярных наблюдений за одним и тем
же гнездом.
Каждая дорога берет свое начало в определенной точке гнезда. Начальные точки
дорог рассредоточены обычно довольно равномерно по периметру гнездового купола.
Логично предположить, что муравьи, действующие на различных дорогах,
придерживаются и в гнезде секторов*, прилегающих к этим дорогам.
Экспериментальная проверка на гнезде обыкновенного лесного муравья,
произведенная нами осенью 1969 г. и летом 1970 г. в Приокско-террасном
заповеднике, подтвердила такое предположение.
Фуражиры на дорогах подопытного гнезда были помечены групповыми метками.
Ночью, когда все фуражиры собрались в куполе, семью усыпили эфиром. Далее был
послойно разобран купол и зафиксировано местонахождение каждого меченого
муравья. Оказалось, что рабочие, связанные с определенной дорогой,
сосредоточиваются в прилегающей к этой дороге части гнезда. Единственным местом,
где рабочие из разных колонн свободно смешиваются, оказался внутренний конус
муравейника. Муравьи-строители также придерживаются своих секторов на
поверхности купола. Привязанность муравьев к своим секторам гнезда оказалась
очень прочной. В ходе разборки муравейника весь строительный материал купола и
оказавшиеся в нем муравьи были полностью перемешаны. Когда разборка гнезда и
регистрация муравьев были закончены, этот материай вместе с начавшими
просыпаться муравьями собрали на прежнее место и придали ему форму,
приблизительно соответствующую доопытной. И в этих условиях фуражиры и строители
нашли свои секторы муравейника.
Еще нагляднее связь колонн с определенной частью гнезда у видов, сооружающих
секционные гнезда, о которых говорилось выше. Там действует четкий принцип:
каждой секции гнезда соответствует колонна. Пространственная разобщенность
колонн сохраняется и на территории. Это вытекает уже из постоянства на
территории кормовых дорог муравейника или кратеров - выходов из секций на
поверхность (рис. 4, 5). Каждая колонна имеет самостоятельный выход или группу
выходов на кормовой участок. На территории характер отношений колонн проявляется
прежде всего во взаимодействии потоков фуражиров разных дорог.
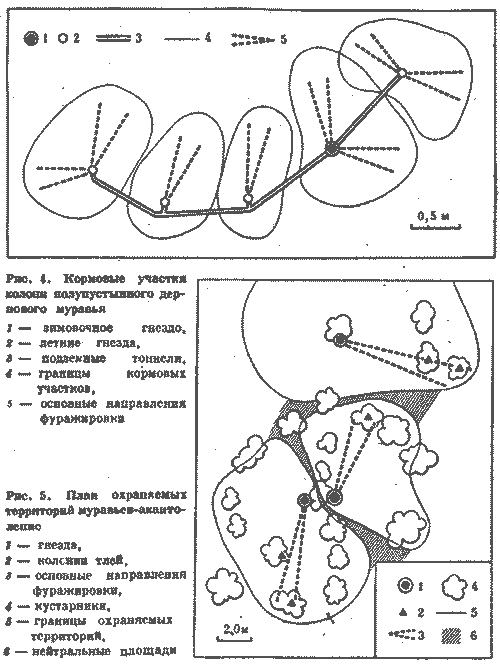
С этой целью в июне 1974 г. был поставлен специальный опыт на средней по
размерам семье обыкновенного лесного муравья (диаметр купола - 0,9 м, высота
гнезда - 0,6 м). Семью составляли четыре колонны примерно одинаковой мощности, о
чем можно судить по сопоставимым размерам дорог (17-20 м) и интенсивности
движения на них (рис. 6). После мочения на всех дорогах 2 тыс. фуражиров в зоне
дороги II была установлена непреодолимая для муравьев металлическая стенка,
вынуждающая поток с дороги II выйти на соседнюю дорогу I. Протяженность стенки в
противоположную от дороги I сторону, к югу, превышала 15 м. Через несколько
часов поток II обошел стенку с севера и вышел на полотно дороги I.
Можно было бы предполагать, что произойдет слияние двух потоков муравьев.
Однако это не случилось. Войдя в вынужденное соприкосновение с потоком II, поток
I отодвинулся от него к северу на 20 см. Миновав препятствие, потоки сразу же
расходились, попадая в собственные русла. Такое положение сохранялось 12 дней,
после чего стенка еще трижды наращивалась, пересекая каждый раз дорогу I. При
этом происходили сопряженные перемещения обоих потоков к северу с сохранением
установившегося между ними расстояния в 20 см. Опыт длился месяц, после чего
стенка была убрана, и уже на следующий день оба потока вернулись в исходное
положение.
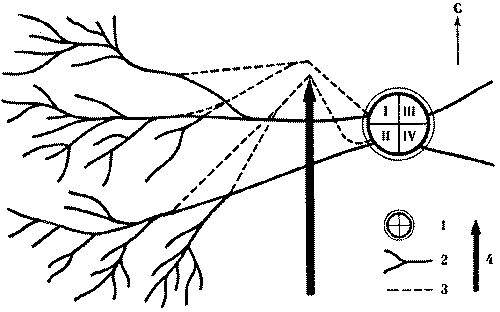
Рис. 6. Потоки фуражиров рыжего лесного муравья разных колонн «отталкиваются» при перемещении дорог.
I, II, III, IV - номера секторов и дорог гнезда; 1 -
муравейник, 2 - положение дорог до и после опыта, 3 - положение
дорог при максимальном смещении во время опыта, 4 - стенка
Этот опыт показал, что существует упругость дорожной сети муравейника, как бы
натянутой между постоянными точками гнезда и кормового участка. Потоки фуражиров
не смешиваются, они отталкиваются друг от друга. В этом выражается
обособленность и целостность колонн и даже противостояние их на участке.
Обособленность колонн на территории сохраняется у рыжих лесных муравьев и в
других частях кормового участка* семьи, где контактируют уже не потоки, а группы
муравьев. Территория каждой колонны охраняется и отделена от охраняемых
территорий* других колонн нейтральными пространствами* - зонами, не посещаемыми
муравьями (так же, как отделены охраняемые территории самостоятельных
муравейников).
Кормовые участки колонн стабильны и неприкосновенны и у видов, не имеющих
дорог. У обитающих в пустыне Каракум зерноядных пустынных дерновых муравьев к
каждой секции примыкает свой обособленный кормовой участок. В ряде случаев он
отделен от других нейтральным пространством, как и у всех зерноядных муравьев.
Сбор семян производится не ежедневно, а по мере их созревания. А так как каждая
колонна собирает семена самостоятельно, то обычна ситуация, когда на поверхности
работают фуражиры лишь части колонн, а выходы из остальных секций закрыты.
Кормовые участки закрытых секций остаются в этот день не использованными ни
фуражирами из других секций данного гнезда, ни другими семьями (рис. 7).
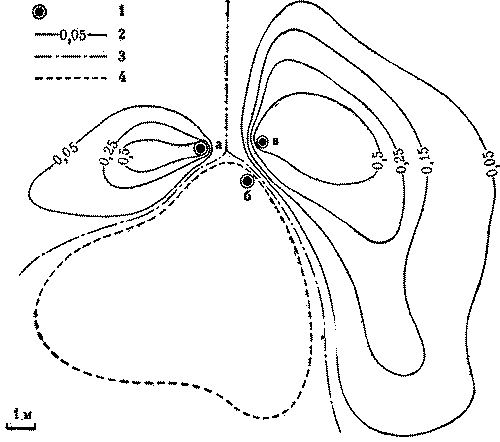
Рис.7. Использование кормового участка трехсекционного гнезда
дернового пустынного муравья при работе двух секций 1 - кратеры секций
а, б а в, г - изолинии динамической плотности особей на участках
работающих секций а и б (особь/дм2мин), 3- границы кормовых
участков секций, 4 - границы охраняемого участка неработающей секции б
Понятно, что только пространственной обособленности колонн в гнезде и на
участке еще недостаточно для существования в муравейнике группировок особей.
Пространственная обособленность должна сочетаться с такими важными
характеристиками, как достаточно глубокая информативная изоляция и
функциональная автономность колонн друг от друга. Естественно, что ни одно из
перечисленных проявлений автономности колонн в рамках семьи не может быть
полным, так как в этом случае семья просто перестала бы существовать как единое
целое.
Информация может передаваться из колонны в колонну различными путями:
посредством обмена пищей, различных сигналов (акустических, киноптических,
химических, тактильных)*; обмена особями (личинками, куколками, взрослыми
муравьями); распространения в гнезде специфического запаха, в результате чего
возникает и поддерживается особый запах муравейника - гнездовой запах*.
Передача различных сигналов носят прежде всего оперативный характер, в то
время как обмен пищей и особями дает информацию, накапливающуюся в семье и
приводящую к изменению поведения со значительной задержкой. Специфический запах
семьи служит как бы эталоном при опознании своих (т. е. из своего гнезда) и
чужих муравьев в гнезде и на участке.
Изучая с помощью изотопов пути распространения пищи в семьях рыжих лесных
муравьев, Р.Шовен установил, что пища, добываемая муравьями на одной из дорог,
не передается в другие секторы гнезда, т. е., как только в семье возникают
колонны, единый пищевой поток, охватывавший ранее всех членов семьи, распадается
на несколько изолированных друг от друга потоков, каждый из которых ограничен
пределами одной колонны. Такая пищевая, или трофическая, изоляция колонн имеет
очень важные для дальнейшего развития муравейника последствия.
Трофическая изоляция колонн и их пространственная обособленность усиливаются
почти полным отсутствием передачи сведений из колонны в колонну о том, что
происходит на кормовом участке. Это было проведено рядом опытов. Один из них
заключался в следующем. У одиночного муравейника с помощью ловушек в течение дня
отлавливали всех фуражиров с одной из дорог, интенсивность движения на
подопытной дороге при этом непрерывно снижалась и через несколько часов упала до
нуля в результате полной выборки всех внегнездовых рабочих из данной колонны.
Казалось бы, для муравейника отлов тысяч фуражиров равнозначен гибели массы
муравьев на участке одной из колонн, что означает серьезную угрозу для всей
семьи.
Однако бедственное положение на одной из дорог не отразилось на скорости и
интенсивности движения и поведении муравьев на остальных дорогах. Там фуражиры
работали в своем обычном ритме.
Такая же картина наблюдалась и на поверхности муравейника, в контрольных
секторах купола. При изъятии всех фуражиров с одной из дорог у нескольких
муравейников, отличающихся по размерам и числу дорог, заметная реакция в
остальных колоннах отсутствовала. Все происходящее на кормовом участке одной
колонны оставалось ее частным делом и совершенно не интересовало соседей,
обитающих в том же гнезде и входящих в состав одной семьи.
Абсолютен ли этот тип изоляции? Конечно, нет. Но вместе с тем настолько
глубок, что мы можем проделать и более бесцеремонные действия, не вызвав тревоги
у соседей. Например, надев резиновые перчатки, отлавливаем с поверхности одного
из секторов купола всех находящихся там муравьев. Здесь, правда, появляется
опасность, что муравьи из соседних секторов заметят резкое движение руки и
устремятся навстречу надвигающейся угрозе. Однако достаточно нехитрого
приспособления, чтобы этого избежать. В нижней части купола и на гнездовом валу,
по границам подопытного сектора, устанавливаем невысокие стенки (5-6 см
высотой). Отлов муравьев производим только между стенками. Непосредственной
реакции муравьев-наблюдателей из соседних колонн на пути сборщика и боевые позы
защитников гнезда из подопытной колонны уже нет. Теперь, если не делать слишком
резких движений, можно отловить всех внегнездовых муравьев одной из колонн без
опасений, что будет захвачено сколько-нибудь значительное число особей из
соседних колонн. Муравьи подопытной колонны реагируют на исчезновение собратьев
и как бы дают сигнал, когда такая выборка закончена: они быстро закрывают все
гнездовые выходы. Сектор как бы вымирает. А рядом в соседних секторах все идет
своим чередом: бдительные наблюдатели патрулируют поверхность купола, строители
укладывают в покровный слой гнезда очередные хвоинки, непрерывным потоком идут в
гнездо и вновь выходят из него потоки фуражиров. Судьба забаррикадировавшейся в
своем секторе колонны никого из них не беспокоит.
Из всего изложенного можно сделать вывод, что колонны, обитающие в одном
гнезде, а у многих видов имеющие даже общую крышу над головой, действуют
фактически обособленно друг от друга. Но очевидно, что такое положение попросту
нереально, ибо тогда непонятно, почему они все-таки обитают в одном гнезде и что
их там удерживает.
Вопрос об автономности и обособленности колонн не возник бы, если бы ее
состав не позволял выполнять в пределах одной колонны все основные функции,
которые свойственны одинарной семье (т. е. семье, не подразделенной на колонны).
В каждой колонне имеются все характерные для вида функциональные группы муравьев
(и внутригнездовых и внегнездовьгх), выращивается молодь. Полное укомплектование
колонны муравьями разных профессий и позволяет ей обособиться.
| Вложение | Размер |
|---|---|
| ant5.gif | 14.75 КБ |
| ant6.gif | 4.97 КБ |
| ant7.gif | 6.37 КБ |
{kind=link}
{kind=link}
{kind=link}
